Infant demographics and SARS-CoV-2 antibody prevalence
This longitudinal analysis included a total of 100,318 newborn screening DBS collected from 50,036 infants between November 2019 and November 2021 who met the inclusion criteria for repeat specimens. Infants with repeat specimens are a select group of infants with an initial sample collected within 4 days of birth as described in detail in the methods section. Data analyses were performed on various groups of infants from this study. See Fig. S2 for a diagram of cohorts and analyses performed.
To evaluate the characteristics of infants with repeat specimens, we compared infants with two specimens (n = 30,344) or more than two specimens (n = 19,692) to a larger subset of infants with a single specimen (n = 377,587) (Fig. 1). As expected, infants with either two or more than two specimens were more likely to be premature (27.3% and 68.3%, respectively) and from a multiple birth (7.3% and 17.9%, respectively) compared to infants with a single specimen (4.8% premature, 2.2% multiple birth). Infants with repeat specimens were also more likely to be of low (18.9% and 41.3%, respectively) and very low (3.3% and 19.8%, respectively) birthweight (BW) compared to infants with single specimens (4.2% low BW and 0.2% very low BW). In addition, infants with two and more than two specimens were more likely to be male (54.3% and 54.8%, respectively) and less likely to be born to individuals 20–30 years old (42.5% and 41.9%, respectively) than those with a single specimen (50.3% male, 45.1% born to individuals 20–30 years old) (Fig. 1).

Demographic characteristics of infants born in NYS between November 2019 and November 2021 with single, two or more than two specimens. Mean percentage (and 95% CI) of infants with single, two or more than two specimens per (a) gestational age (wk), (b) birthweight (g), (c) sex, (d) maternal age (y), and (e) multiple birth status.
To evaluate SARS-CoV-2 antibody reactivity in infants with single or repeat specimens, we tested DBS for IgG antibodies to SARS-CoV-2 S and N antigens and evaluated demographic characteristics of initially reactive infants and the timing of antibody development11. Infants were grouped based on antibody reactivity at birth and, for nonreactive samples, subsequent antibody results. Overall reactivity for S or N SARS-CoV-2 IgG antibodies at birth was significantly higher in infants with a single specimen (n = 80,551; 21.3%, 95% CI 21.2–1.5%) than in infants with repeat specimens (n = 9,611; 19.2%, CI 18.9–19.6%) (Table 1). Of infants with repeat specimens, 630 (1.3%) were nonreactive for IgG antibodies at birth and developed either S or N antibodies in subsequent specimens (Table 1).
Antibody reactivity in infants with single or repeat specimens was analyzed to identify associations with demographic characteristics of the infant and/or mother using logistic regression. When compared to infants that were IgG nonreactive at birth, infants that were antibody reactive at birth were less likely to be from a multiple birth (odds ratio [OR] 0.88, 95% confidence interval [CI] 0.84–0.92, p < 0.001), less likely to be born with low birthweight (OR 0.95, CI 0.91–0.99, p = 0.01), less likely to have repeat DBS specimens (OR 0.93, CI 0.90–0.96, p < 0.001), and less likely to have both repeat specimens and very low birthweight (OR: 0.67, CI 0.55–0.83, p < 0.001). Antibody reactive infants were more likely to be born to individuals > 30 years old (OR 1.15, CI 1.10–1.21, p < 0.001) (Table 2).
To evaluate the patterns of SARS-CoV-2 antibody reactivity, we plotted percent reactivity in the initial birth DBS of infants with single or repeat specimens over the 25-month period beginning November 2019 (Fig. 2). All infants born before March 2020 were nonreactive for both S and N SARS-CoV-2 IgG antibodies. The percentage of infants with reactivity to S or N SARS-CoV-2 IgG increased from March 2020 through June 2020, increased again from November 2020 through April 2021, and continued to increase from July 2021 through November 2021, corresponding to the increase in infections and COVID-19 vaccinations among individuals giving birth during this period (Fig. 2). Throughout the study period, initial antibody reactivity among infants with repeat specimens was lower than in infants with single specimens, but patterns of reactivity were similar.
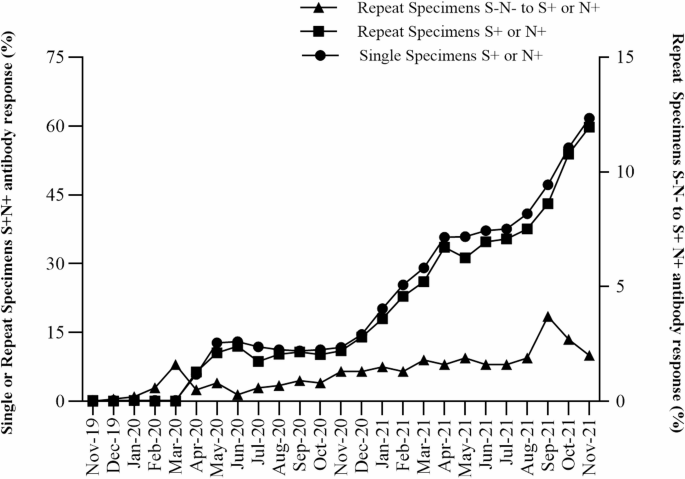
SARS-CoV-2 spike (S) and nucleocapsid (N) antibody reactivity in NYS infants with single or repeat specimens between November 2019 and November 2021. Percentage of infants with single or repeat specimens born S+ or N+ SARS-CoV-2 antibody reactive (S+N+) (left y-axis, circles or squares, respectively) by month of birth. Percentage of infants with repeat specimens born SARS-CoV-2 antibody non-reactive who became S+ or N+ antibody reactive in a post-birth specimen (S–N– to S+ or N+) (right y-axis, triangles) by midpoint month between initial and last specimen.
Infants with repeat collections that were nonreactive at birth and developed S or N antibody reactivity in subsequent samples were analyzed to identify periods of seroconversion between November 2019 and November 2021. Seroconversion in the infant post-birth is likely due to the infant becoming infected with SARS-CoV-2 and developing their own antibodies, since infants who were noted to have received blood transfusions were excluded from the study. The estimated seroconversion date was calculated as the midpoint between the collection dates of the birth and the final specimen. The first estimated seroconversion, where an infant developed SARS-CoV-2 antibody reactivity post-birth, occurred in an infant who was non-reactive at birth in February 2020 and seroconverted by March 2020, a period which coincided with the first reported cases of COVID-19 in NYS. The percentage of infants who seroconverted post-birth rose in September 2021 and declined by November 2021 (Fig. 2).
Longitudinal analysis of infant SARS-CoV-2 antibodies
Next, we evaluated longitudinal changes in antibody reactivity in infants with more than two DBS. We chose this population because the average time between the first and last specimen was greater in infants with more than two specimens (22.7 ± 0.2 days), compared to infants with two specimens (11.4 ± 0.1 days). This would allow more time to elapse between specimens, increasing our ability to detect changes in antibody reactivity. However, infants with more than two specimens were demographically different from infants with a single specimen, especially with regards to birthweight and gestational age (Fig. 1). As such, we examined the effect of these factors on longitudinal changes in antibody reactivity in subsequent analyses.
Maternally-derived SARS-CoV-2 antibody decay in infants
Infants passively acquire IgG maternal antibodies during pregnancy and titers of these maternal antibodies decay over time. We analyzed SARS-CoV-2 antibody decay in infants with more than 2 samples using linear mixed-effects modeling, including 2,400 infants with S reactive IgG antibodies and 579 infants with N reactive IgG antibodies passively acquired. A selective analysis was used to exclude infants who may have seroconverted in utero and whose antibody levels increased post-birth, by restricting the dataset to infants who were initially antibody reactive (MFI index > 1) at birth and whose antibody levels had declined in the last specimen collected from that infant (Fig. S1). In this population, antibody levels declined in a non-linear process, exemplified by a rapid decay in the first 30 days followed by a slower decay (Fig. 3). Half-lives were estimated for infants born S (and N) IgG antibody reactive for each of these periods (0–30 days after birth, > 30 days after birth), including infants with initially high, intermediate, and low levels of antibody (Fig. 3, Table 3, Table S1, Table S2). Half-life is the time required for the concentration of maternal IgG antibodies to decrease by half. Overall, S and N IgG half-life estimates were significantly lower 0–30 days after birth (22.8 and 22.1 days, respectively) compared to > 30 days after birth (37.5 and 36.6 days, respectively), confirming our observation of a faster rate of decay for the first 30 days followed by a slower decay after 30 days. This pattern (longer half-lives after 30 days) was consistent at low, intermediate, and high levels of S IgG and N IgG, although not all categories were significantly longer. Within each period (0–30 and > 30 days), half-lives were not significantly different between low, intermediate, and high initial antibody levels. Therefore, initial high reactivity was expected to persist longer than low and intermediate reactivity, due to higher initial MFI index values (Fig. 3) with similar rates of decay.

Natural log of MFI index values for infants born SARS-CoV-2 IgG antibody reactive for (a–d) spike (S) and (e–h) nucleocapsid (N). Antibody decay in the total population (a,e) or in subgroups of infants with initial high (b,f), intermediate (c,g) and low (d,h) levels of SARS-CoV-2 IgG antibodies. Slopes for antibody reactive infants were allowed to change after 30 days to provide a linear approximation to the non-linear decay process.
Since our dataset includes a high proportion of infants who were born premature or with low or very low birthweight, we determined if these characteristics impacted half-life estimates. Although not significant, half-life estimates of N IgG > 30 days after birth were lower in premature infants than in infants with normal gestational age (95% CIs barely overlapped, Table 3).
Alternative models can be useful to analyze non-linear patterns of antibody decay. We used logistic regression to analyze antibody decay in 2,400 infants reactive for S IgG and 579 infants reactive for N IgG based on the mean probability, for an infant that was previously reactive, that their next sample would be non-reactive (i.e. seroreversion). We estimated a mean IgG seroreversion probability of 56 days for S and 28 days for N (95% CI 50–62 for S and 95% CI 24–33 days for N, respectively) (Fig. 4, Table S3).

Probability of non-reactive SARS-CoV-2 (a) spike (S) IgG and (b) nucleocapsid (N) IgG antibodies in subsequent specimens collected from infants born in NYS. Black dots indicate reactive (1’s) and non-reactive (0’s) infants. Blue dots show the percent of infants within each bin that were reactive. Hosmer–Lemeshow Goodness of Fit test p = 0.49 for S IgG and p = 0.381 for N IgG. Sample sizes were 2,400 infants for S IgG and 579 infants for N IgG. Shading indicates a 95% confidence interval of the mean probability estimate. The probability of a reactive repeat specimen dropped below 50% after 56 days for S IgG (95% CI 50–62 days) and was 28 days for N IgG (95% CI 24–33 days).
Infant SARS-CoV-2 antibody seroconversion
Of 19,692 infants with more than two specimens, seroconversion was detected in 240 infants who were nonreactive for SARS-CoV-2 IgG antibodies in the initial specimen but became reactive in subsequent collections. Twenty-three infants seroconverted for both S and N IgG, 205 for S IgG only, and 12 for N IgG only.
IgG in newborns is indicative of passive acquired immunity24, whereas detection of IgM and IgA post-birth suggests active immunity. Therefore, to confirm that infants had seroconverted, we retested a select group of 34 IgG seroconverting infants for the presence of SARS-CoV-2 IgM and IgA antibodies (see Supplement S3). Five patterns of seroconverting antibody isotypes were observed (see Fig. 5 for representative examples): synchronous S or N IgG, IgM and IgA (8 infants), synchronous S or N IgG and IgA (10 infants), synchronous S or N IgG and IgM (3 infants), S IgG only (13 infants), and infants born S IgG reactive that subsequently seroconverted for either IgA or IgM (9 infants). Of infants who seroconverted for IgG post-birth, 20.6% (and 29.4%) of infants seroconverted to S (and N) IgM, and 52.9% (and 20.6%) seroconverted to S (and N) IgA. We also used a modeling approach to evaluate antibody seroconversion rates in this population (see Supplement S2). Detection of IgA and IgM in several infants confirmed seroconversion for SARS-CoV-2 in this population.

Antibody levels (ln(MFI Index)) against SARS-CoV-2 spike (S) and nucleocapsid (N) in individual infants with 3 or more DBS who seroconverted between initial and subsequent samples (a.-h., 34 infants) or were initially seropositive for IgG S and seroconverted for IgA or IgM (i.-j., 9 infants). Horizontal dotted line indicates reactive cutoff (ln(MFI Index) = 0). Infant identifiers are given at the top of each panel for reference.
We also tested 144 infants that were reactive for S IgG at birth to identify if any had detectable IgM and IgA at birth, which could indicate in utero infection (see Supplement S3). One infant was reactive for N IgM (ID 6695101) at birth. No IgM or IgA was detected in the birth specimen of the remaining 143 infants that were IgG positive at birth. We also determined if these initially IgG reactive infants had later seroconverted for IgM or IgA, a finding that would indicate an infection post-birth. Of the 9 infants that seroconverted for IgA or IgM but were reactive for S IgG at birth, one (ID 1905147) seroconverted > 4 weeks post-birth and became reactive for N IgG, N and S IgA, and N and S IgM. Of the remaining eight, five seroconverted for N IgA and three seroconverted for N IgM. Interestingly, S IgG appeared to decline in four infants while N IgA or N IgM increased. This unique pattern may be explained by maternal derived S IgG antibody decay that masked an infant derived S IgG antibody response.




